Plasmodesmata De brug tussen plantencellen
 Share
Share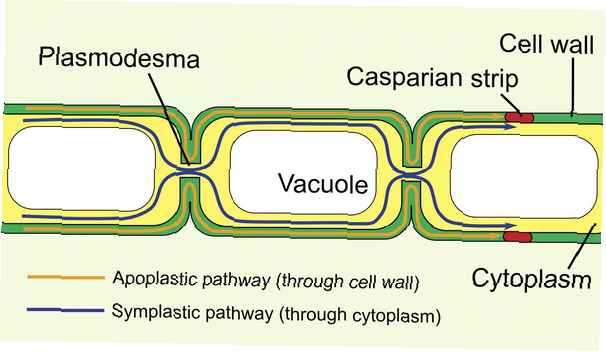
Plasmodesmata is een dun kanaal door plantencellen waarmee ze kunnen communiceren.
Plantencellen verschillen op veel manieren van dierlijke cellen, zowel in termen van sommige van hun interne organellen als in het feit dat plantencellen celwanden hebben, waar dierlijke cellen dat niet hebben. De twee celtypen verschillen ook in de manier waarop ze met elkaar communiceren en hoe ze moleculen transloceren.
Wat zijn Plasmodesmata?
Plasmodesmata (enkelvoudige vorm: plasmodesma) zijn intercellulaire organellen die alleen in planten- en algencellen worden gevonden. (De dierlijke cel "equivalent" wordt de gap junction genoemd.)
De plasmodesmata bestaat uit poriën of kanalen die tussen individuele plantencellen liggen en verbinden de symplastische ruimte in de plant. Ze kunnen ook worden aangeduid als "bruggen" tussen twee plantencellen.
De plasmodesmata scheiden de buitenste celmembranen van de plantencellen. De werkelijke luchtruimte die de cellen scheidt, wordt de desmotubule genoemd.
De desmotubule bezit een star membraan dat de lengte van het plasmodesma doorloopt. Cytoplasma ligt tussen het celmembraan en de desmotubule. Het gehele plasmodesma is bedekt met het gladde endoplasmatisch reticulum van de verbonden cellen.
Plasmodesmata vormen zich tijdens celdeling van plantontwikkeling. Ze vormen zich wanneer delen van het gladde endoplasmatisch reticulum van de oudercellen vast komen te zitten in de nieuw gevormde plantencelwand.
Primaire plasmodesmata worden gevormd terwijl ook de celwand en het endoplasmatisch reticulum worden gevormd; secundaire plasmodesmata worden daarna gevormd. Secundaire plasmodesmata zijn complexer en kunnen verschillende functionele eigenschappen hebben in termen van de grootte en aard van de moleculen die er doorheen kunnen.
Activiteit en functie
Plasmodesmata spelen rollen bij zowel cellulaire communicatie als bij de translocatie van moleculen. Plantencellen moeten samenwerken als onderdeel van een meercellig organisme (de plant); met andere woorden, de individuele cellen moeten werken om het algemeen belang te bevorderen.
Daarom is communicatie tussen cellen cruciaal voor het overleven van planten. Het probleem met de plantencellen is de harde, stijve celwand. Het is moeilijk voor grotere moleculen om de celwand binnen te dringen, en daarom zijn plasmodesmata noodzakelijk.
De plasmodesmata verbinden weefselcellen met elkaar, zodat ze functioneel belang hebben voor weefselgroei en -ontwikkeling. Onderzoekers verduidelijkten in 2009 dat de ontwikkeling en het ontwerp van belangrijke organen afhankelijk waren van het transport van transcriptiefactoren (eiwitten die RNA helpen omzetten in DNA) door de plasmodesmata.
Plasmodesmata werd eerder gedacht als passieve poriën waardoor voedingsstoffen en water werden verplaatst, maar nu is bekend dat er sprake is van actieve dynamiek.
Actinestructuren bleken transcriptiefactoren te helpen verplaatsen en zelfs plantenvirussen door het plasmodesma te verplaatsen. Het exacte mechanisme van hoe de plasmodesmata het transport van voedingsstoffen regelen, is niet goed begrepen, maar het is bekend dat sommige moleculen ervoor kunnen zorgen dat de plasmodesma-kanalen breder worden geopend.
Fluorescerende sondes hielpen bij het vinden dat de gemiddelde breedte van de plasmodesmale ruimte ongeveer 3-4 nanometer is. Dit kan echter variëren tussen plantensoorten en zelfs celtypen. De plasmodesmata kunnen zelfs in staat zijn om hun dimensies naar buiten te veranderen zodat grotere moleculen kunnen worden getransporteerd.
Plantenvirussen kunnen mogelijk door plasmodesmata bewegen, wat problematisch kan zijn voor de plant, omdat de virussen rond kunnen reizen en de hele plant kunnen infecteren. De virussen kunnen zelfs de grootte van het plasmodesma manipuleren zodat grotere virale deeltjes erdoorheen kunnen bewegen.
Onderzoekers geloven dat het suikermolecuul dat het mechanisme regelt voor het sluiten van de plasmodesmale porie callose is. Als reactie op een trigger zoals een pathogeeninvaller wordt callose afgezet in de celwand rond de plasmodesmale porie en sluit de porie.
Het gen dat de opdracht geeft om callose te synthetiseren en af te zetten, wordt CalS3 genoemd. Daarom is het waarschijnlijk dat de plasmodesmatadichtheid de geïnduceerde weerstandsreactie op pathogeenaanval in planten kan beïnvloeden.
Dit idee werd verduidelijkt toen werd ontdekt dat een eiwit, genaamd PDLP5 (plasmodesmata-gelokaliseerd eiwit 5), de productie van salicylzuur veroorzaakt, wat de afweerreactie tegen plantpathogene bacteriële aanval verbetert.
Onderzoeksgeschiedenis
In 1897 merkte Eduard Tangl de aanwezigheid van de plasmodesmata op in het symplasm, maar het was pas in 1901 toen Eduard Strasburger ze plasmodesmata noemde.
Door de introductie van de elektronenmicroscoop konden de plasmodesmata natuurlijk nader worden bestudeerd. In de jaren tachtig konden wetenschappers de beweging van moleculen door de plasmodesmata bestuderen met behulp van fluorescerende sondes. Onze kennis van de structuur en functie van plasmodesmata blijft echter rudimentair en er moet meer onderzoek worden verricht voordat alles volledig wordt begrepen.
Verder onderzoek werd lang belemmerd omdat plasmodesmata zo nauw geassocieerd zijn met de celwand. Wetenschappers hebben geprobeerd de celwand te verwijderen om de chemische structuur van de plasmodesmata te karakteriseren. In 2011 werd dit bereikt en veel receptoreiwitten werden gevonden en gekarakteriseerd.
